 |
Прочитано: 86% |
«««Назад | Оглавление | Каталог библиотеки | Далее»»»
|
Прочитано: 86% |
В главах 4 и 5 мы рассмотрели физиологические явления тормозной доминанты и имитативного рефлекса и показали, что их встреча в высшей нервной деятельности семейства троглодитид должна была породить высоко развитое явление интердикции. Наибольшую ступень интердикции мы назвали генерализованной интердикцией: имитационное провоцирование некоторого одного действия парализует возможность каких бы то ни было других действий (очевидно, за исключением функционирования автономной нервной системы) и это состояние парализованности, вероятно, может продолжаться долго - даже после прекращения действия данного имитатогенного агента.
Тем самым высшую форму интердикции можно было бы считать низшей формой суггестии: это уже не торможение лишь того или иного отдельного действия, но навязывание некоего состояния, допустим, типа каталепсии. Однако таков лишь зачаток суггестии, ибо под ней (под "внушением") понимается возможность навязывать многообразные и в пределе даже любые действия. Последнее предполагает возможность их обозначать. Между этими предельными рубежами умещается (развивается) явление суггестии. Нас прежде всего интересуют его первые шаги.
Надо напомнить еще раз коренной тезис, что вначале, в истоке, вторая сигнальная система находилась к первой сигнальной системе в полном функциональном и биологическом антагонизме. Перед нашим умственным взором отнюдь не "добрые дикари", которые добровольно подавляют в себе вожделения и потребности для блага другого: они обращаются друг к другу средствами инфлюации, к каковым принадлежит и суггестия, для того чтобы подавлять у другого биологически полезную тому информацию, идущую по первой сигнальной системе, и заменять ее побуждениями, полезными себе. Это явление инфлюации, в том числе суггестии, не имеет никакого отношения к гносеологии. Сколько обвинений в идеализме и мистицизме было обрушено на вывод Леви-Брюля о "прелогическом мышлении", тогда как это явление действительно существовало на заре истории и проявлялось впоследствии, но только оно не было "мышлением": оно было подавлением первосигнального (еще единственно верного тогда) способа отражения окружающей среды и системой принудительного воздействия на поведение друг друга. Да, оно принадлежит не к гносеологии, а к онтологии. Это только взаимодействие особей. Тут нет отношения субъекта к объекту, а есть лишь отношение организма к организму. "Прелогическая стадия" ничуть не угрожает логике: тут логике еще решительно нечего было делать. Мы убедимся, что и антилогика в самом деле гораздо раньше объявилась в этой сфере отношений между индивидами, чем логика в сфере человеческого познания. Но о генезисе логики мы будем говорить только в конце этой главы (и, тем самым, этой книги).
Развитие второй сигнальной системы у людей ни в коем случае не было следствием разрастания общего объема головного мозга по сравнению с объемом (весом) тела и прямо никак не связано с этим процессом энцефализации (церебрализации) в филогении троглодитид. Во-первых, в биологической эволюции вообще налицо тенденция увеличения мозга (независимо от размеров тела), и по этому показателю прямоходящие высшие приматы, включая человека, вовсе не оказываются специфичными: степень энцефализации увеличивается у всех ископаемых млекопитающих. По степени развития неокортекса ("новой коры"), т.е. по степени "неоэнцефализации", можно выделить весь отряд приматов из общей родословной млекопитающих, но невозможно отдифференцировать собственно человеческую линию. Во-вторых, разрастание объема головного мозга в эволюции семейства троглодитид было прямым морфологическим следствием прямохождения и прямым биохимическим следствием плотоядения, т.е. повышенного усвоения протеина ; эти два фактора влекли за собой широкий размах индивидуальных вариаций объема мозга, иначе говоря, делали структурно и гистологически возможными колебания в сторону повышенной массы мозга, а естественный отбор закреплял эти отклонения, несомненно, потому, что они были биологически выгодны. По Боне, это разрастание мозга происходило в том же темпе, как и другие морфологические трансформации млекопитающих в плейстоцене. Но у Homo sapiens средний размер и вес головного мозга не возрастал и не возрастает сравнительно с поздними палеоантропами. При этом размах индивидуальных вариаций данного признака у Homo sapiens весьма увеличился сравнительно с палеоантропами, нередко встречается объем мозга, значительно превышающий среднюю величину, но ничто не закрепляет этих отклонений: они биологически нейтральны и средняя величина остается неподвижной для черепов любого времени с верхнего палеолита до наших дней. Между тем речевая функция мозга в корне отличает неоантропа от палеоантропа.
Как видно, социальность и разум человека никак прямо не коррелированы с тотальной величиной его головного мозга.
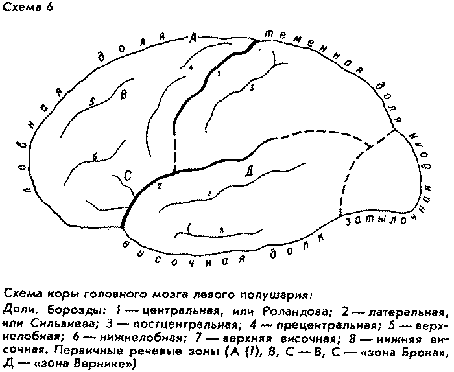
Зато корреляция начинает проступать, когда измерению подвергаются по отдельности длиннотный, широтный и высотный диаметры роста головного мозга в филогенетической цепи: шимпанзе - австралопитек - археоантроп - палеоантроп - неоантроп. До неоантропов наиболее интенсивно увеличивался тотальный длиннотный размер (за счет роста задних областей); напротив, только мозг неоантропов дает интенсивный сдвиг высотного диаметра: в лобной и теменной долях эпицентры роста перемещаются из нижних отделов в верхние, происходит, как мы уже отмечали, усиленный рост префронтальной области, а также поднятие и выравнивание поверхности мозга в его своде.
«««Назад | Оглавление | Каталог библиотеки | Далее»»»
|
| ||||||||
|
|
|
|
|
|
|
|||